/Image2482.gif) 地質時代と示準化石 |
/Image2481.gif) 示準化石 ウィキペディア(HP/2012/11)による『示準化石』から |
|
|
最終更新日:2017年2月11日
| 全般 | 動物 | 植物 |
|
その他 |
|
リンク| 化石の定義| 示準化石| 生命の誕生| 最古の化石| 絶滅| |
三葉虫| アンモナイト| |
植物| |
フズリナ(紡錘虫)|
放散虫| |
種の定義| 生物の大分類| ストロマトライト| |
|
化石(Fossil)とは、過去の生物(古生物、Prehistoric
Life)の痕跡(Trace)を言う。化石によって、地質年代(Geological Age)が決められてきた。 地質年代における時代の境界(Boundary)は、その当時の主要な生物の絶滅(大量絶滅、Mass Extinction)を意味する。従って、過去の地球史におけるさまざまなイベント(Event)の発生の結果を反映している。 生命(Life)の起源(Origin)と進化(Evolution)の過程を明らかにするためにも、化石の研究は不可欠である。 |
| リンク |
| 全般 | 動物 | 植物 |
|
その他 |
|
リンク| 化石|生痕化石|微化石| 古生物学| 生命の起源|生物の進化| 絶滅| |
化石動物| エディアカラ動物群| バージェス動物群| |
化石植物| 珪化木| |
フズリナ(紡錘虫)|
放散虫| コノドント| |
ストロマトライト| カンブリア爆発| 石灰岩をつくる化石| べリンガーの化石| その他 |
【エディアカラ動物群】〔約6~5.5億年前〕(Ediacara)
| 化石の定義 |
| 示準化石(Index fossil) |
|
地質時代と示準化石 |
|
示準化石 ウィキペディア(HP/2012/11)による『示準化石』から |
| 生命の誕生 |
|
生命発生の条件
生命の特徴
中村(1994)による〔『生命進化40億年の風景』(29-33p、44-45p)から〕【見る→】 |
| 最古の化石 |
| 絶滅 |
/Image2351.gif) The circled peaks on this graph from the paper are the mass extinctions while the dotted lines mark each 27 million years Firthによる『Life on Earth wiped out every 27 million years (but we've got about 16 million years until the next one)』(2010/7) |
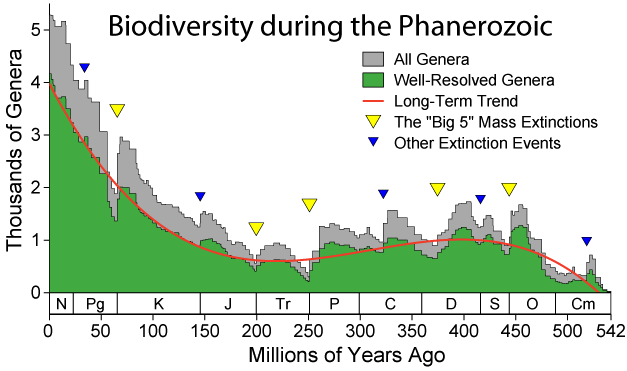 〔Wikipedia, the free encyclopediaのImage:Extinction Intensity.png(Extinction event)から〕 Genera:Genus(属)の複数形。 |
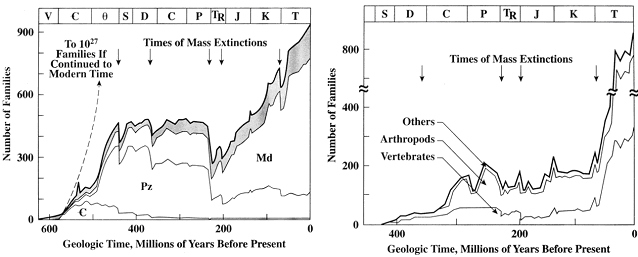 Figure 18.4. Number of biological families of [left] shelly marine fauna and [right] nonmarine animals over time. Dashed line extrapolates to later times the increse in number of families in the early Cambrian in the absence of extinctions. Arrows show times of mass extinctions. On the nonmarine graph, vertebrates and arthropods are broken out as well. Note that animals did not appear in nonmarine sediments, and hence were not on land, until almost 200 million years after the start of the Phanerozoic. The top portion of the graph labels the geologic periods using traditional symbols. From Milne et al. (1985). 〔University of WyomingのPaul Heller氏による『GEOLOGY 1200: Historical Geology』の中の『Lecture 11: ORIGIN OF LIFE』から〕 |
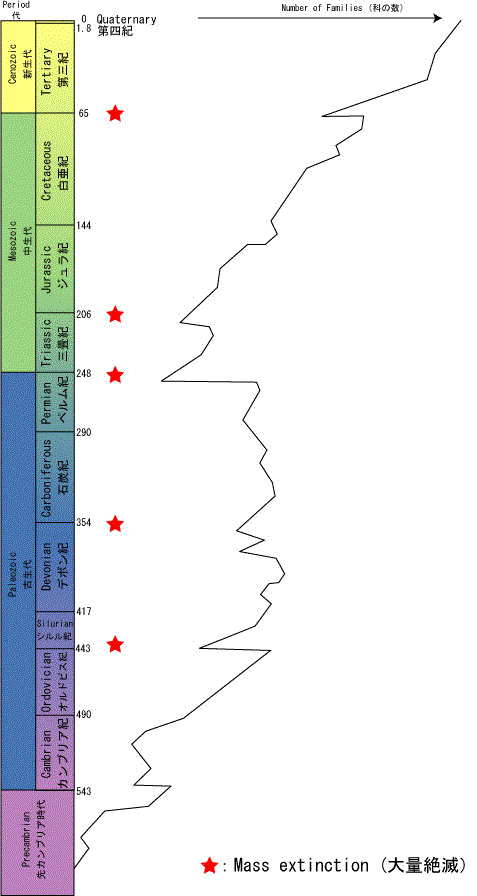 図7 顕生代を通じての海洋無脊椎動物の多様性の変化 (多様性のデータは,Sepkoski, 1984, Benton, 2001を元にした.地質時間スケールは,GSA, 1999を元にしています.) |
/Image2349.gif) 図1:5回あった顕生代の大量絶滅 /Image2350.gif) 図4:地球内の要因が絡まって起きた古生代/中生代境界の大量絶滅 磯崎(2005)による『大量絶滅 生物進化の加速装置』から |
| 表2 地史を通じてみられる主要な絶滅と環境変動。①~⑪が古生代以降の11大絶滅、下線がジャブロンスキーの5大絶滅。 | |||||
|
|
|
|
|
|
|
| ⑪中新世中期 | (15) | 軟体動物 | 気候寒冷化 | 25.0(0.43) | 12 |
| ⑩始新世後期 | (36.5) | 軟体動物 | 海水準低下 | 45.8(0.50) | 15 |
| ⑨白亜紀末 | (65) | 浮遊性有孔虫、斧足類、アンモナイト、ベレムナイト、鳥盤類、竜盤類 |
海水準低下+ 気候寒冷化+ 隕石衝突 |
66.3(1.74) | 90 |
| ⑧白亜紀後期CT | (91) | アンモナイト、斧足類 | 無酸素事変 | 18.9(0.93) | 36 |
| ⑦白亜紀前期AA | (107) | 斧足類 | 無酸素事変 | 12.0(0.22) | 18 |
| ⑥ジュラ紀末 | (135) | 斧足類、アンモナイト、竜脚類恐竜、剣竜 |
海水準低下+ 気候変動 |
19.5(1.09) | 30 |
| ⑤ジュラ紀前期PT | (188) | 斧足類 | 無酸素事変 | 15.2(0.86) | 17 |
| ④三畳紀後期 | (205) | コノドントアンモナイト(セラタイト亜目)、腕足類、腹足類、斧足類、両生類(迷歯類)、哺乳類型爬虫類、槽歯類 | 海水準低下 | 38.6(1.94) | 36 |
| ③ペルム紀末 | (250) | サンゴ、フズリナ、腕足類、海百合、コケムシ、アンモナイト(ゴニアタイト亜目)、哺乳類型爬虫類 |
海水準低下+ 気候寒冷化 |
D:52.5(5.61) G: - (7.12) |
81 154 |
| 石炭紀末 | (290) | - | |||
| ②デボン紀後期FF | (360) | 造礁性生物(サンゴ、海綿)、腕足類、三葉虫、アンモナイト、コノドント、板皮類 |
海水準低下+ 氷河発達 |
21**、50* | |
| シルル紀末 | (410) | - | 海水準低下 | ||
| ①オルドビス紀末 | (438) | 造礁性生物(サンゴ、海綿)、筆石、コケムシ、腕足類、オウムガイ、三葉虫 |
氷河発達+ 海水準低下 |
22** | |
| カンブリア紀後期 | (510) | 三葉虫、コノドント、腕足類 |
海水準低下?+ 気候寒冷化? |
15~20* | |
| 先カンブリア時代後期 | (650) | 藻類(アクリタークス) | 氷河発達 | ||
|
古生代以降の11大絶滅は、ジャブロンスキー、スタンレー、ラウプ、セプコスキーのいう大量絶滅をカウントしたもの。先カンブリア時代については情報が乏しく、まだ十分に議論されていないのでカウントから除いた。ペルム紀以降の絶滅率はラウプとセプコスキー(1984)による。( )内の絶滅率は、セプコスキーとラウプ(1986)による(絶滅科数)÷(全科数)を100万年あたりで表示した値。科数は絶滅事変のあった期の間の絶滅科数。すべて海生動物に基づく。 *:セプコスキー(1986)による海生動物の属の絶滅率(%)。 **:セプコスキー(1982)による海生動物の科の絶滅率(%)。地質時代は紀単位で表示してあるが、紀末の絶滅率は、その紀の最後の期の絶滅率である。 CT:セノマニアン期とチューロニアン期の境界、AA:アプチアン期とアルビアン期の境界、PT:プリーンスバキアン期とトアルシアン期の境界、FF:フラスニアン期とファメニアン期の境界、D:ズルフィアン期、G:グアダルピアン期。 |
|||||
| 三葉虫 |
/Image2484.gif) 三葉虫綱 参照資料 A Guide to the Orders of Trilobites三葉虫の分類 『切手に見る世界の昆虫』(HP/2012/11)の『三葉虫綱』から |
/Image2486.gif) In the image above, trilobites (1) live among many species that are not normally preserved. A typical Cambrian outcrop might produce only trilobites, brachiopods (2), mollusks (3), and crinoids (4). That is a tiny fraction of the full Cambrian biota, better represented by the roster of the Burgess Shale Cambrian Konservat-Lagerstatten. That community includes sponges Vauxia (5), Hazelia (6), and Eifellia (7); brachipods Nisusia (2); priapulid worms Ottoia (8); trilobites Olenoides (1); other arthropods such as Sidneyia (9), Leanchoilia (10), Marella (11), Canadaspis (12), Helmetia (13), Burgessia (14), Tegopelte (15), Naraoia (16), Waptia (17), Sanctacaris (18), and Odaraia (19); lobopods Hallucigenia (20) and Aysheaia (21); mollusks Scenella (3); echinoderms Echmatocrinus (4); and chordates Pikaia (22); among other oddities, including Haplophrentis (23), Opabinia (24), Dinomischus (25), Wiwaxia (26), Amiskwia (27), and Anomalocaris (28). /Image2485.gif) Systematic Relationships and Chronological Extent of the Trilobite Orders Sam Gon III(HP/2012/11)による『A Guide to the Orders of Trilobites』の『Trilobite Systematic Relationships』から |
/Image2487.gif) 頭部(cephalon)、胸部(thorax)、尾部(pygidium)。 ウィキペディア(HP/2012/11)による『三葉虫』から |
| アンモナイト |
/Image2483.gif) アンモナイトが生息していた時期と進化 アンモライト研究所(HP/2012/11)による『アンモナイトについて』から |
|
2-3.Page (1996)の説  |
| 植物 |
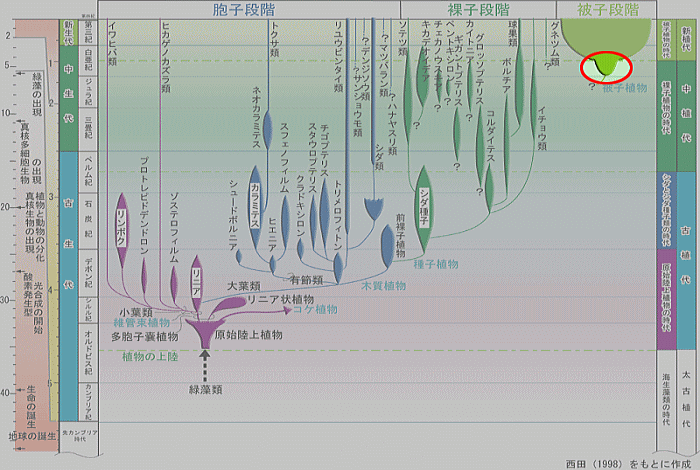 〔大阪市立自然史博物館の『化石からたどる植物の進化』の中の『-系統樹- 被子植物』から〕 |
| フズリナ(紡錘虫:石灰質の殻をもつ) |
/Image1235.gif) Fusulinida |
/Image1236.gif) Fusulinid limestone, Upper Pennsylvanian; Elk County, Kansas. Field of view is 3.9 cm wide. Link at source to view of entire slab. |
| Wikipedia(HP/2014/11)による『Fusulinida』から | |
/Image1240.gif) コレヂャ・ストーン(2005/7)による『紡錘虫の部』から |
/Image1237.gif) This cutaway view of a fusulinid test shows the complex structure of these single-celled organisms. The prominent line on the outside of the text, the antetheca, was the growing surface, where new chambers were added (drawing by Roger B. Williams, KU Paleontological Institute). Brosius(2005/9)による『Fossil Fusulinids』から |
| 放散虫(ラジオラリア:主に珪酸質の骨格をもつ) |
/Image1241.gif) 放散虫 "Cyrtoidea" from Ernst Haeckel's Kunstformen der Natur, 1904 ウィキペディア(HP/2014/11)による『放散虫』から |
/Image1243.gif) 図 2 代表的な古生代放散虫の骨格のスケッチ. |
|
/Image1244.gif) 図 5 古生代放散虫の科の産出年代範囲. バーの幅は属の数を示す. |
/Image1242.gif) 図 7 古生代放散虫の属の数と絶滅イベ ン ト. 矢印は7回 の絶滅イベ ン トを示す. |
| 梅田(2002)による『古生代放散虫の分類と消長史-7回の絶滅事件-』から | |
| 種の定義 |
中村(1994)による〔『生命進化40億年の風景』(6-11p)から〕【見る→】 |
||||||||||||||||||
| 生物の大分類 |
|
| ストロマトライト |
| ストロマトライト(stromatolite)は、藍藻(シアノバクテリア)類の死骸と泥粒などによって作られる層状の構造をもつ岩石のことである。特に、内部の断面が層状になっているものを指す。 | |
/Image2606.gif) こぶのように見える先カンブリア紀のストロマトライトの化石。Siyeh層中にあるものを撮影(米国モンタナ州グレイシャー国立公園) |
/Image2607.gif) ストロマトライト〔西オーストラリア州・セティス湖(Lake Thetis)〕 |
| ウィキペディア(HP/2015/10)による『ストロマトライト』から | |
/Image1233.gif) 図 1重要なストロマトライト産出地. 現世(海成):1 シャーク湾(オーストラリア),2 リーストッキング島(バハマ);現世(非海成):3 クーロン・ラグーン(オーストラリア),4 グリーン湖(アメリカ合衆国);顕生代:5 ブント砂岩(ドイツ・トリアス紀);先カンブリア時代:6 ビッタースプリングス層(オーストラリア・原生代),7 ピルバラ地塊(オーストラリア・太古代),8 ロックネスト層(カナダ・原生代),9 ポンゴラ地塊(南アフリカ共和国・太古代). |
/Image1232.gif) 図 2 代表的な先カンブリア時代のストロマトライト形態のスケッチ. 1-5 層状ストロマトライト:1,2 Stratifera Koroliuk,3 Tyssagaetes Vlasov,4 Irregularia Koroliuk,5 Malginella Komar and Semikhatov;6-10 ノジュール状ストロマトライト:6-8 ドーム型:6 Paniscollenia Koroliuk,7 Collenia Koroliuk,8 Plicatina Raaben,9,10“キャベツ型”:9 Cryptozoon(sensu Pia),10 Cryptophyton Raaben and Komar;11-13 柱状無分枝ストロマトライト:11 Colonella Komar,12 Conusella Golovanov,13 Conophyton Maslov;14 Yakutophyton Schapovalova;15 Songjiella Cao et al.;16-19 微小ストロマトライト:(スケールは 1 cm) 16 Asperia Semikhatov,17 ochmecolumella Liang and Zhang,18 Minicolumella Raaben,19 Vetella Krylov;20-30 柱状有分枝ストロマトライト;20 Gymnosolen Steinmann,21 Boxonia Koroliuk,22-24 ツングーシア科:22 Tungussia Semikhatov,23 Anabaria Komar,24 Baicalia Krylov,25 Kussiella Krylov,26-30 イントリカティア科:26 Kussoidella Semikhatov,27 Sundia Butin,28 Pilbaria Walter,29 Confunda Semikhatov,30 Kanpuria Raaben.Semikhatov and Raaben(2000)を改変. |
/Image1231.gif) 図 4 シアノバクテリアの活動によるGebeleinのストロマトライト生長モデル.昼間はシアノバクテリア(Schizothrix calcicola)の上位方向への移動と砕屑物の固着が起こりストロマトライトが生長する(a,b).夜間はシアノバクテリア(Oscillatoria submembranacea)の水平方向への移動と砕屑物の固定が顕著に起こる(c).Gebelein(1969)を改変. |
|
/Image1230.gif) 図 5 現世のストロマトライト形成場(サブカ)でみられる,ストロマトライトの形態変化とそれを形成するシアノバクテリアの種.形態によって種が異なる.Golubic(1976)を改変. |
/Image1229.gif) 図8 先カンブリア時代のストロマトライト礁復元図.ストロマトライトの形態は環境に応じて変化する.Grotzinger(1990)を改変. |
| 山本・磯崎(2013)による『ストロマトライト研究の歴史と今後の展望』から | |
 Living stromatolites at Shark Bay, Western Australia, showing the laminated structure of a single column and the microscopic cyanobacteria that construct them. Each cyanobacterial filament is about 100th of a millimetre in width. These organisms are rarely preserved in fossil stromatolites, although some have been discovered in chert (silica-rich rock) at other localities in the Pilbara that are about the same age as the Trendall locality. 〔DoIRによる『Geological Survey of Western Australia』の『Discover Geology』の中の『Stromatolites』から〕 |
 オーストラリア西海岸のハメリンプール(Hamelin Pool)にある現生ストロマトライト 〔Science Circle Hokkaido JapanによるWisdom96の中の『Hamelin Pool』から〕 オーストラリアのハメリンプール〔Hamelin Pool Marine Nature Reserve:⇒シャーク湾(Shark Bay)〕はストロマトライト(Stromatolite)の名所。ストロマトライトはシアノバクテリア〔ラン藻、藍藻(Cyanobacteria、Blue-Green Algae、Blue-Green Bacteria、Cyanophyta)〕などの光合成(Photosynthesis)に伴う分泌物によって形成された炭酸塩岩(Carbonate rock)である。35億年前頃から現在まで存在する。地球大気中の酸素(Oxygen Gas)は、これらの生物により生み出されたと信じられている。 |
/Image1238.gif)
/Image1239.gif)
{kind=link}