/Image2641.gif) 【図1】地球上への微生物の出現・進化 横田による『微生物分類同定講座』から |
/Image2640.gif) 微生物の分類学上の位置付け エフジーエー・ラボラトリーズ(株)(HP/2015/12)による『微生物とは』から |
|
|
最終更新日:2018年9月2日
| 全般 | 細菌 | 原生生物 | 藻類 | その他 |
| リンク|
微生物|病原微生物| |
有孔虫|
放散虫| |
珪藻| | ウィルス| |
|
肉眼(Naked Human Eye)で判別できないような微小な(Microscopic)生物(Organism)を微生物(Microorganism、Micro Organism、Micro-organism、Microbe)と呼ぶ。 その中には多様な生物種(Biological Species)が含まれる。その多くを原核生物(Prokaryote)が占める。 プランクトン(Plankton)という用語は浮遊生物(Drifting Organisms)を指すが、微生物と同様の意味で使われることも多い。 |
| リンク |
| 全般 | 細菌 | 原生生物 | 藻類 | その他 |
|
リンク| 微生物|病原微生物| 組織・機関| 講義ノート| 用語集| |
原核生物| 細菌(真正細菌)| 古細菌| マイコプラズム|リケッチア|クラミジア| |
原生生物| 有孔虫| 放散虫| |
藻類| 珪藻| 渦鞭毛藻| 円石藻| |
ウイルス| プランクトン| ミドリムシ(ユーグレナ)| ベントス| デトリタス| 下水道| その他 |
|
(1)微生物は森を育てる (2)水の中の微生物 (3)からだの中の微生物 (4)水田や畑をつくる微生物 (5)微生物が食べ物をつくる (6)極限に生きる微生物 (7)恐ろしい微生物 (8)資源をつくる微生物 (9)薬をつくる微生物 (10)環境を守る微生物 (11)微生物が地球をつくった (12)微生物の多様性 |
【原核生物】(prokaryotes、Prokaryota)
| 微生物 |
【図1】地球上への微生物の出現・進化 横田による『微生物分類同定講座』から |
|
微生物の分類学上の位置付け エフジーエー・ラボラトリーズ(株)(HP/2015/12)による『微生物とは』から |
|
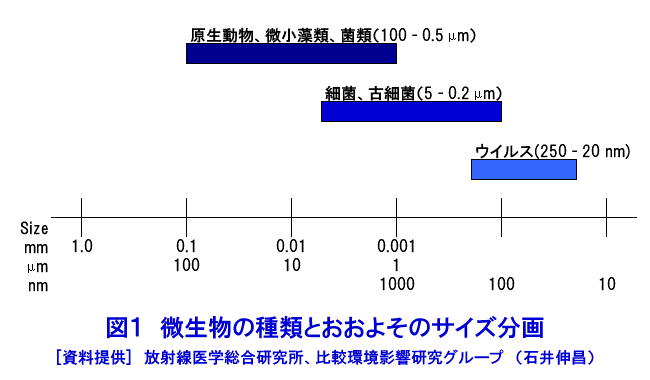
微生物とは、肉眼でその存在が判別できず、顕微鏡などによって観察できる程度以下の大きさの生物を指す。微生物を研究する学問分野を微生物学と言う。 ウィキペディア(HP/2014/2)による『微生物』から |
/Image2642.gif) 原子力百科事典ATOMICA(2005/9)による『土壌微生物と放射性核種の環境挙動とのかかわり』から |
| 病原微生物 |
細菌・マイコプラズマ・リケッチア・クラミジア・ウイルスの特徴 (有)オオワダによる花野井薬局の感染症のチョコット知識の『感染症のチョコット知識? インフルエンザウイルスによる感染症』(HP/2018/8/29)から |
病原微生物の分類と特徴 |
病原微生物の大きさ |
|
|
| (株)アトラスによる感染症.comの『病原微生物の分類と特徴について』(2017/7/3)から | |
(株)アトラス感染症.com運営事務局による『病原微生物の分類と主な特徴』(HP/2015/12)から |
| 細菌(真正細菌) |
 真正細菌の基本的な構造 細胞膜の外側には細胞壁(この画像ではそのさらに外側に莢膜)がある。細胞内小器官は存在せず内容物は混ざっている。 |
|
|
|
| ウィキペディアによる『真正細菌』(HP/2018/8/27)から | |
/Image2649.gif) 全生物を描いた系統樹の例。左側のバクテリアが真正細菌に相当する。この系統樹では、グラム陰性菌とグラム陽性菌が系統的なまとまりを示している。 ウィキペディア(HP/2015/12)による『真正細菌』から |
/Image2643.gif) 【図1】16S rRNA遺伝子の塩基配列に基づく細菌および古細菌の代表菌種の分子系統樹、ならびに対応する門レベルの系統群 Bergey's Manual of Systematic Bacteriology Second Edition(2001)に採用されている古細菌2門および真正細菌23門を示した。2007年現在、新たにGammatimonadetes、LentisphaeraeおよびTenericutesの3門が真正細菌の系統群として提唱されている。赤線で示された菌種は超好熱菌であることを示す。 |
真正細菌 |
| 横田による『微生物分類同定講座』から | |
/Image2650.gif) 森本(2010/7)による『第75話 微生物と人類との関わり~微生物には不可能はない!~』から |
| 有孔虫 |
化石有孔虫(Foraminifera)でよく知られているのは、古生代(石炭紀~ペルム紀)のフズリナ(fusulina、Fusulinida)〔広義=紡錘虫類(fusulinid)、狭義=紡錘虫類のうちのフズリナ属〕と新生代第三紀の貨幣石(かへいせき、Nummulite、ヌムリテス)である。【リンクはウィキペディア】
/Image2488.gif) Figure : Maximum likelihood phylogeny of Rhizaria inferred from SSU rRNA gene sequences using the GTR+G+I model of evolution. Molecular Systematics Group(HP/2012/11)による『Tree of Rhizaria』から Rhizaria=リザリア、Cercozoa=ケルコゾア、Foraminifera=有孔虫、Haplosporidia⇒アセトスポラ〔略胞子虫(Haplosporidia)類とパラミクサ(Paramyxea)類の2群からなる〕=Ascetosporea、Gromia=グロミア、Phytomyxea=ネコブカビ類、Radiolaria=放散虫。 【リンクはウィキペディア】 |

国際海洋環境情報センターによる浮遊性有孔虫データベースから 現在の浮遊性有孔虫の例(グロビゲリナ、Globigerina)。約0.1ミリメートル程度の大きさ。糸状の仮足は剥がれてしまっていて見えない。 |
| 放散虫 |
 Figure 6. Schematic diagram of the mechanisms that can distort the sedimentary imprint of the planktonic pattern of fossilizable microplankton in general, and of polycystine radiolarians in particular (see text for detailed explanation). From Boltovskoy 1995.
Figure 6. Schematic diagram of the mechanisms that can distort the sedimentary imprint of the planktonic pattern of fossilizable microplankton in general, and of polycystine radiolarians in particular (see text for detailed explanation). From Boltovskoy 1995.
Boltovskoy(1998/8/1)による『Classification and distribution of South Atlantic Recent polycystine Radiolaria』から |
|
|
| Radiolaria.org(HP/2016/10/12)による『Silica distribution in the World Ocean: In the water column and in the bottom sediments』から | |
| 珪藻 |
|
* Tréguer,Paul, Nelson,David M., Van Bennekom,Aleido J., DeMaster,David J., Leynaert, Aude and Quéguiner,Bernard(1995): The Silica Balance in the World Ocean: A Reestimate. Science, 268 (5209), 375–379. Bibcode:1995Sci...268..375T. doi:10.1126/science.268.5209.375. PMID 17746543. Retrieved 2013-11-13. |
|
* Egge,J.K. and Aksnes,D.L.(1992): Silicate as regulating nutrient in phytoplankton competition. Mar Ecol. Prog. Ser., 83, 281–289. doi:10.3354/meps083281. |
| Wikipedia(HP/2016/10/12)による『Diatom』から | |
The Trustrees of Indiana University(HP/2016/10/12)による『Introduction to Oceanography Marine Sediments Unit B』から |
| ウィルス |
中屋敷均による『生物と非生物の境界、ウイルスとは何か』(2017/7/5)から |
ウィキペディア(HP/2015/12)による『Template:ボルティモア分類』から |
||||||||||||||||||||||||||||||
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
/Image2647.gif) ウイルスの基本構造(上)エンベロープを持たないウイルス (下)エンベロープを持つウイルス |
/Image2648.gif) 細胞(左)とウイルス(右)の増殖様式 |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ウィキペディア(HP/2015/12)による『ウイルス』から | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
/Image2644.gif) 図2:ウイルスと真正細菌の比較。ヘルペスウイルスは従来から知られているウイルスの代表として、それ以外のウイルスは巨大ウイルスのグループとしてあげた。標準的な真正細菌の代表として大腸菌を、小型の真正細菌の代表としてマイコプラズマをあげた。 |
|
/Image2645.gif) 図3:巨大ウイルスも含めた生きもののグループ分け。由来が分からない孤児遺伝子の多さから、巨大ウイルスも生きものと並ぶ新たなグループではないかと考えている。 |
/Image2646.gif) 図5:ゲノム解析で得られた巨大ウイルスの系統樹。赤文字は既にゲノムが決まっている巨大ウイルスの系統、青文字は海水のメタゲノム解析から存在が推測される系統。 |
| 緒方(2014)による『巨大ウイルスから見える新たな生物界の姿』から | |
| プランクトン |
漂泳生物のうち、遊泳できるものをネクトン(nekton、遊泳生物:魚類など、動物のみ。幼生の間のみ一時プランクトンとなるものがいる。)、遊泳できないものをプランクトン(plankton、浮遊生物)と呼ぶ。漂泳生物に対し、水底で生活するものをベントス(benthos、底生生物)と呼ぶ。【リンクはウィキペディア】
(株)日本海洋生物研究所(HP/2015/2/2)による『プランクトン』から |
||||||||||||||||||||||||
/Image2478.gif) 滋賀県琵琶湖環境科学研究センター(HP/2012/11)による『琵琶湖の主なプランクトン』から |
 〔浜崎恒二氏による『海洋微生物生態学入門』から〕 |
|
図1:約7,000種が知られている動物プランクトンのごく一部の面々。 (1)(2)はカイアシ類。ボートのオール(かい)のような脚(あし)をもつためこの名がある。 動物プランクトンの中では数・量・種数ともに圧倒的に大きい。「海のお米」とも呼ばれ、ほとんどの魚はカイアシ類を食べて大きくなる。大きさは普通数mm以下。 (3)は「海の妖精」とも呼ばれるクリオネ。実は貝の仲間。 (4)も泳ぐ巻き貝のカメガイ。 (5)は肉食性のヤムシ類。カイアシ類をおもな餌とする。 (6)はクラゲノミ類、その名のとおりクラゲなどに寄生する。 (7)はオキアミ類。 「動物プランクトン」は水の中に棲み、泳ぐ力があまり大きくない動物を全てひっくるめた呼び名であり、きわめて多様な動物群を含むため、形や生活も実に多様性に富んでいる。カイアシ類、ヤムシ類、オキアミ類など数量の多いグループでは生活史や生態系の中での役割が良く知られている種もあるが、分布や生態がほとんど分かっていない種も多い。さらには、未だ発見されていない膨大な数の種がいると思われる。 【リンクはウィキペディア】 |
/Image2479.gif) |
| 東京大学大気海洋研究所海洋生態系動態部門浮遊生物分野による『動物プランクトンの多様性を探る - 全海洋動物プランクトンセンサス(Census of Marine Zooplankton: CMarZ)-』(2009/2)から | |
/Image2480.gif) 原島(1993)による『人工衛星可視域データのグローバルマッピングによる広域海洋環境変動に関する研究』から |
| ミドリムシ(ユーグレナ) |
 ユーグレナの細胞構造 1:細胞核、2:葉緑体、3:パラミロン顆粒、4:収縮胞、5:基底小体、6:陥入部(apical pocket)、7:短鞭毛(後鞭毛)、8:感光部(副鞭毛体)、9:眼点、10:長鞭毛(前鞭毛) ウィキペディアによる『ユーグレナ藻』(HP/2018/8/25)から |
 ミドリムシ細胞の模式図 |
|
| 丸山真一朗・野崎久義による『むかしむかし、ミドリムシは紅かった?』(2011/4/19)から | |
| バイオマス |
堀越・二井(2003)による『土壌微生物生態学』から |
||||||||||||||||||||||||||||||||||||
/Image2616.gif)
/Image2617.gif)







 Map showing annual production of biogenic silica in the plankton of the world oceans (g/m2/yr)
Map showing annual production of biogenic silica in the plankton of the world oceans (g/m2/yr)




 Figure 4.19 from your text summarizes the distribution of marine pelagic and neritic sediments
Figure 4.19 from your text summarizes the distribution of marine pelagic and neritic sediments



{kind=link}